ことの始まり
国立障害者リハビリテーションセンター研究所に赴任し、研究の主要テーマの一つとして挙げたのが、網膜色素変性症(RP)の新規原因遺伝子候補の探索です。それまで知られている原因遺伝子のほとんどは視細胞で発現しているものでしたが、視細胞の働きを助けている網膜色素上皮細胞 (retinal pigment epithelium, RPE)で発現している遺伝子の中にも原因となる遺伝子があると考え探索することにしました。
経緯
新しく開発したベクターキャッピング法を用いて、ヒトRPEの細胞株であるARPE-19から完全長cDNAライブラリーを作製し、このライブラリーから任意に選んだ24,000クローンについて、5’端の部分塩基配列を決定しました(K11-2)。得られたクローンの中からRPの原因遺伝子候補として取り上げたのがアリルスルファターゼI (ARSI)です。その理由は、アリルスルファターゼ遺伝子の変異がムコ多糖症のようなリソソーム蓄積症を引き起こし、RPに似た網膜変性を伴う場合があることと、ESTデータに基づくARSI遺伝子の発現プロフィールを求めたところ眼で多く発現していることが示されたからです。
我々のヒト完全長cDNAコレクションの中にアリルスルファターゼA(ARSA)とアリルスルファターゼの活性化に必要なスルファターゼ修飾因子(SUMF1)の完全長cDNAも含まれていたので、ARSAは活性測定の対照として、SUMF1は活性化因子として用いました。まずARSIのC端にFLAGタグを融合させたタンパク質をARPE-19で発現させ、免疫蛍光染色法により細胞内局在を求めました。ついでARPE-19細胞でARSIとSUMF1を発現し、基質として4-methylumbelliferyl sulfate(4-MUS)を用いて発現産物のアリルスルファターゼ活性を測定しました。
さらに、ロドプシン遺伝子の変異スクリーニング(K07-1)に用いた68名のRP患者のゲノムDNAを用いて、第5染色体上にあるARSI遺伝子の変異スクリーニングを行いました。
結果
ARSIタンパク質の細胞内局在を調べたところ、ERに局在が認められましたが、予想に反してリソソームには認められませんでした。ウェスタンブロッティングにより発現産物の解析を行ったところ、ARSAとSUMF1は細胞抽出物と培地に存在するが、ARSIはほとんど細胞ライセートの不溶画分にあることがわかりました。また、生産量もARSAやSUMF1に比べ著しく少ないことが示されました。
細胞抽出物の可溶画分を用い4-MUSを基質としてアリルスルファターゼ活性を測定したところ、活性が認められないどころか内在性のアリルスルファターゼ活性を低下させるという予期せぬ結果が得られました。ARSIの活性化に必要な93番目のCysをSerに変えたARSI変異体を発現した場合やSUMF1を共発現させた場合は、このような低下が認められませんでした。
培地にバンドは認められませんでしたがTCA沈殿物にわずかながらバンドが認められ、弱いながら中性付近に示適pHを有するアリルスルファターゼ活性が認められました。以上の結果からARSIは分泌型のアリルスルファターゼであると推定しました。
本来の目的である変異スクリーニングの結果、RP患者のゲノムDNAのARSI遺伝子領域にRPの原因となるような変異は見つかりませんでした。
余談
我々がARSI cDNAをクローン化したのは2003年であり、本研究を開始した時点でARSIに関する報告はまだありませんでした。2005年Sardielloらはヒトゲノム配列の中からホモロジー検索により、4種類のアリルスルファターゼ遺伝子を見つけ、ARSH、ARSI、ARSJ、ARSKと命名しました(Sardiello et al., 2005)。2006年Obayaはヒト胎児肺のcDNAライブラリーからARSIのcDNAをクローン化し、C端にFLAGタグをつけてHeLa細胞で発現させたところ、生成物は不溶画分に見られ、4-MUSを基質として用いたアリルスルファターゼ活性は検出されませんでした(Obaya, 2006)。また免疫染色でリソソームへの局在は確認できませんでした。
2026年になってGrecco-Machadoらは、ニワトリ胚の軟骨成熟過程でプロテオグリカンの硫酸化が減少する現象に関与する因子としてARSIを同定し、ヒトARSIがリソソームに局在し、コンドロイチン硫酸プロテオグリカンを基質とするアリルスルファターゼであることを見出しました(Grecco-Machado et al, 2026)。プロテインAと融合させることにより、高収量のARSI融合タンパク質を得ることができたようです。
本研究は東洋大学大学院生であった押川未央さんの博士論文の一部になっています。押川さんは膨大な数の実験をこなし、興味ある多くの結果を出してくれました。
疑問点
Q1 ARSIを培養細胞で発現させた場合、ARSAに比べ生産量が少なく、かつ不溶画分にくるのはなぜか。
ARSIとARSAは同じプロモータの発現ベクターを用いているので、転写量に大きな違いはないと考えられます。一つの要因として考えられるのはN端やC端のアミノ酸配列の違いです。特にARSIのC端にARSAやARSBにない46アミノ酸残基からなる配列が存在することは注目に値します。この領域の特徴は連続する8個の酸性アミノ酸残基とそれに続く15個の塩基性アミノ酸に富む領域、そして2個のCysの存在です(図1)。
図1 ARSI、ARSA、ARSBのアミノ酸配列の比較
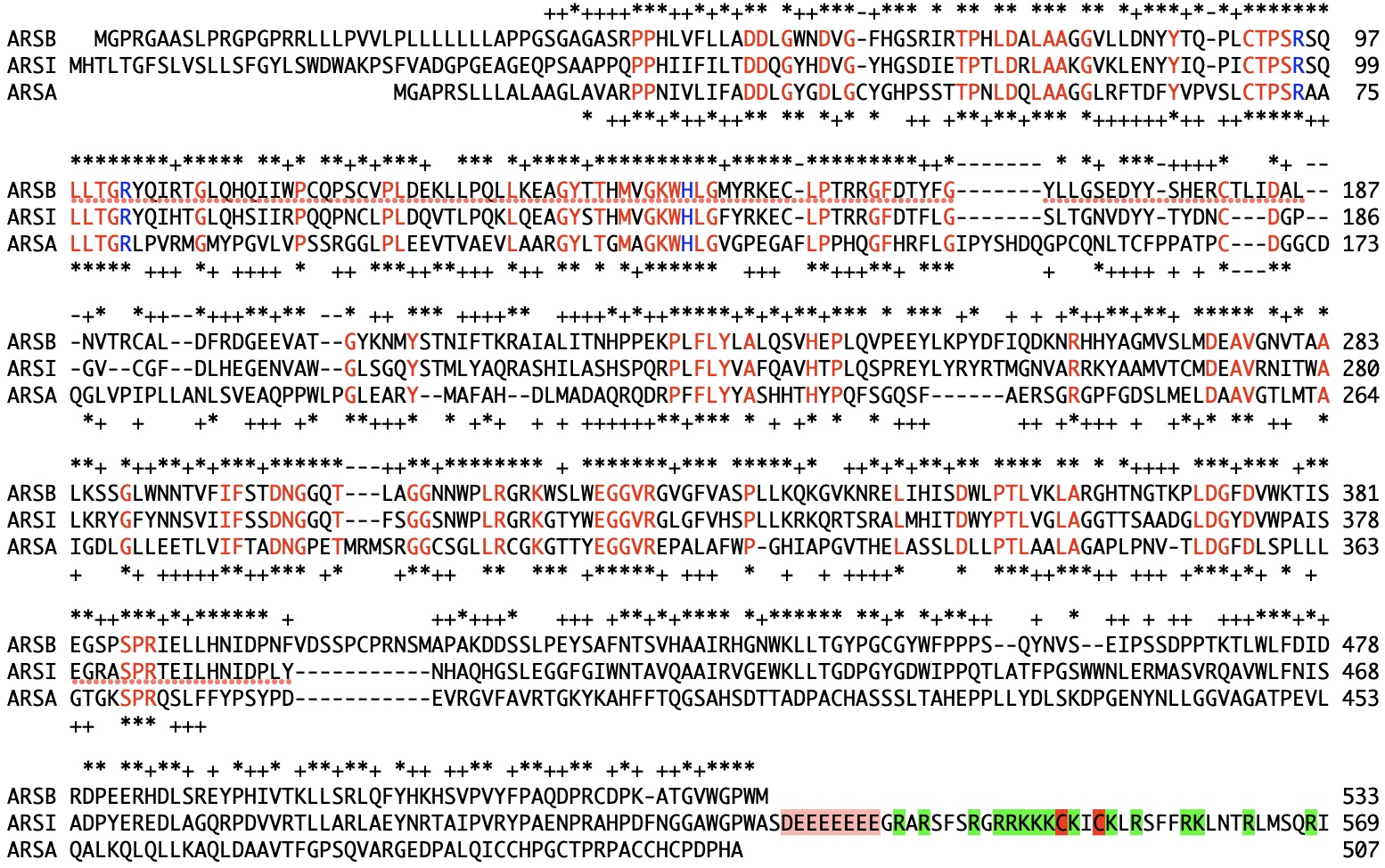
本論文では、このC端46残基を欠失したC端欠失ARSIの生産を試みました。その結果、この組換え体の不溶画分の生産量は増加しましたが、可溶画分にはほとんどバンドが認められませんでした(Mol. Vis., Figure 2D)。したがって、C端の余分な配列は生産量低下に関与しているが、不溶化には関与していないと考えられます。このFigure 2Dで興味深い点は、C端欠失ARSIの場合、不溶画分と可溶画分のいずれにも分解産物と思われる25kDa以下の複数のバンドが見られることです。
Grecco-Machadoらは、ARSIのN端にProtein Aを融合したProtein A-ARSI融合タンパク質をCOS7細胞で分泌生産させています(Grecco-Machadoら、Figure 8B)。ただ、IgG-Sepharoseカラムで精製する前の細胞ライセートや培地のウェスタンブロッティングの結果が載っていないので、不溶化や分解が起こっているかどうかは不明です。ARSIのN端に融合することによって、融合タンパク質が不溶化せずに生産できたとすると、ARSIのN端側の配列が不溶化の原因になっている可能性があります。
{kind=link}
Q2 ARSIの局在がリソソームに認められなかったのはなぜか。
そもそも細胞抽出物の可溶画分にバンドが認められないので、リソソームへの輸送が行われていたとしてもその量が少なく検知できるレベルではなかった可能性があります。Grecco-Machadoらは、ARSIのN端にGFPを融合したGFP-ARSI融合タンパク質をHeLa細胞で生産させリソソームへの局在を見ています(Grecco-Machadoら、Figure 4)。我々はARSIのN端ではなくC端にタグを付加しており、この違いが両者の発現量の違いや不溶化の有無の原因になったのかもしれません。
{kind=link}
Q3 ARSIの網膜色素上皮細胞における生理的役割は何か。
Grecco-Machadoらは、ARSIの生理的基質はコンドロイチン硫酸プロテオグリカンであり、軟骨成熟過程に関与していることを明らかにしました。網膜に軟骨はありませんが、光受容体間マトリックス(IPM: interphotoreceptor matrix)の主要成分がコンドロイチン硫酸プロテオグリカンであることから、網膜色素上皮細胞によって貪食された視細胞の表面に含まれるIPMの分解過程でARSIが機能していると考えられます。
ちなみに我々のヒト完全長cDNAコレクションの中に、ARSJ、SUMF2、光受容体間マトリックス遺伝子IMPG2、5種のコンドロイチン関連遺伝子(CHST11、CHST12、CHST3、CHPF、CHSY1)が含まれています。
Q4 ARSIの活性測定結果をどう解釈するか。
ARSIのアミノ酸配列と最も類似性が高いのはARSBです(図1)。したがって、生理的基質もARSBと同様、コンドロイチン硫酸プロテオグリカンであると想定されました。ただ、当時我々の研究室ではコンドロイチン硫酸を基質とするアッセイ系がなかったので、ARSBの基質となりうる4-MUSを使用しました。コンドロイチン硫酸のアッセイ系があったとしても、ARSIの生産量が少ないので活性測定できたかは疑問です。
生産量が少ないので精製せずに活性測定を行いました。ベクターだけを導入した細胞、活性部位を潰したCys93Ser変異体を発現させた細胞、SUMF1を共発現した細胞を対照として用いました。問題は発現ベクターを入れない細胞でも微量の内在性ARSによる活性があり、細胞抽出液と培地のARS活性のpH依存性が異なることです。しかも、細胞抽出液の場合、ARSIを発現させると他の対照よりも活性が低下するという現象が見られました。Discussionの中でこの現象の解釈を試みましたが、まだ多くの疑問が残ります。
Q5 ARSAやARSIをSUMF1と共発現すると、SUMF1の分泌が認められなくなるのはなぜか。
ARSIのみならずARSAやARSIのCys93Ser変異体でも同様の現象が見られることから、ARSがSUMF1と相互作用し、何らかの形でSUMF1がER内にトラップあるいは分解され分泌できなくなったのではないかと考えられます。これについてはもっと追求したかったのですが、我々の本来の目的はRPの原因遺伝子探索であったので、さらなる追求は行いませんでした。
被引用文献
Google Scholarで検索した結果、被引用数は現時点(2026年)で16となっています。被引用数が少ないのは、ARSIに関する研究がほとんど進展していないからと思われます。ARSIタンパク質に関するものよりは、種々の条件下で発現量が変化する遺伝子の一つとしてARSIが候補に上がってきたという研究が主です。引用されている事柄の内訳を見てみると、ARSIの説明[3、6、14]、ERへの局在と分泌[1、9、10、12、16]、ARPE-19での発現[5、7、8]、ARSIの発現プロフィール[15]、RPとの関連[13]、ARSIの活性[16]に関する引用となっています。ARPE-19の説明[2]や ARSJの説明[4]などARSIとは無関係な事柄で引用されているものもあります。